


| name | Amanita ochraceopallida |
| name status | nomen acceptum |
| author | Contu |
| english name | "Contu's Pallid Ringless Amanita" |
| images |
 1. Amanita ochraceopallida, Sardinia, Italy. |
| intro | The following description is based on the original description by Contu (1998). |
| cap | The cap of A. ochraceopallida is 20 - 100 mm wide, uniformly very pallid with ochraceous brown tint, campanulate-convex, eventually broadly convex, with or without an obtuse umbo, slightly viscid when moist, with a striate margin. The volva is present as white, unchanging warts of varying small sizes. |
| gills | The gills are free, moderately crowded, ventricose, white, with a flesh pink or pale cream tint, with an uneven, slightly flocculose and concolorous edge. |
| stem | The stem is 70 - 130 × 8 - 15 mm, cylindric, narrowing upward, white, with a frost-like concolorous covering which is rarely broken up into whitish concentric bands. No ring is present. The volva is membranous, saccate, persistent, white on the outside, tending to be grayish with yellow-orange spotting. The flesh is white and unchanging. |
| odor/taste | The odor and taste are weak. |
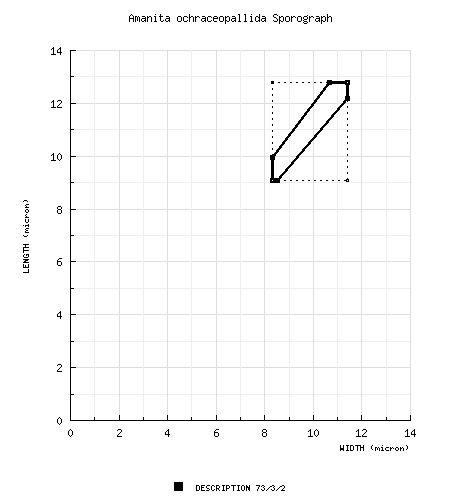
| spores | Spores measured by RET are (9.0-) 9.1 - 12.8 (-16.5) × (8.0-) 8.3 - 11.4 (-13.5) µm and are subglobose to broadly ellipsoid and inamyloid. Clamps are absent at bases of basidia. |
| discussion |
This species was originally described from Sardinia (Italy) in a mountain habitat under plantations of fir (Abies cephalonica). This species is rare and localized on Sardinia. In the notes on two of the paratypes, Dr. Contu states that "this is the sole Amanita species occurring in Abies plantations in Sardinia." Dr. Contu was kind enough to send RET portions of two of the paratypes of this species from which the spore measurements above were taken. Since the spore measurements provided in the original description are exactly the same as those provided here with the extremes of length and width eliminated, it may be that the type materials' spores were not measured. Revision of the type would be useful. Based on the material RET has examined, A. ochraceopallida is closest to the species group listed on the A. crocea (Quél. in Bourd.) Singer ex Singer taxon page.—R. E. Tulloss and L. Possiel |
| brief editors | RET |
| name | Amanita ochraceopallida | ||||||||
| author | Contu. 1998 ["1997"]. Micol. Veg. Medit. 12: 137-139, figs. 1-3, pl. (p. 145 bottom, mislabeled). | ||||||||
| name status | nomen acceptum | ||||||||
| english name | "Contu's Pallid Ringless Amanita" | ||||||||
| MycoBank nos. | 446315 | ||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||
| holotypes | CAG | ||||||||
| revisions | Tulloss, herein. | ||||||||
| intro |
The following text may make multiple use of each data field. The field may contain magenta text presenting data from a type study and/or revision of other original material cited in the protolog of the present taxon. Macroscopic descriptions in magenta are a combination of data from the protolog and additional observations made on the exiccata during revision of the cited original material. The same field may also contain black text, which is data from a revision of the present taxon (including non-type material and/or material not cited in the protolog). Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The following material is derived from the protolog of the present species and from original research of R. E. Tulloss. | ||||||||
| pileus | from protolog: 2-10 cm, not very fleshy, campanulate-convex expanding with age, always obtusely umbonate, slightly lubricous-viscous, of a typical pale yellowish-ochre color, margin concolorous, surface frequently ornate with firm, white creamy veil residues (rarely verrucose). | ||||||||
| peridium | double click in markup mode to edit. | ||||||||
| lamellae | from protolog: ventricose, thick, free, white, pale flesh-colored with age, section concolorous, floccose-crenulate. | ||||||||
| stipe | from protolog: 7-13 × 0.8-1.5 cm, elongated, cylindric, progressively elongated towards the base but never bulbous, white, ornate with a creamy concolorous pruinose material, rarely mottled with whitish bands. Ring absent. Volva of the vaginata type, membranous, long sheathing, persistent, non-friable, white, sometimes tending to gray or with yellow-orange spots. Flesh not very firm, white, unchanging. | ||||||||
| odor/taste | from protolog: taste and odor weak, not very characteristic. | ||||||||
| macrochemical tests |
none described. | ||||||||
| pileipellis |
from protolog: composed of wide interwoven cylindric hyphae 3-10.5 µm, vacuolar pigment; suprepellis is a clear an deep ixocutis of very slender and confusingly erect hyphae. from RET study of paratypes: 105 - 135 μm thick; suprapellis (15-) 30 - 40 μm thick, colorless, gelatinized to partially gelatinized; subpellis 75 - 85 (-120) μm thick, yellow-orange in exsiccata, largely ungelatinized; filamentous undifferentiated 1.7 - 6.0 μm wide, branching, except over disc hyphae predominantly subradially oriented but with some criss-crossing, over disc as rather open lattice, densely packed vertically in both supra- and subpellis; vascular hyphae 3.8 - 11.5 μm wide, plentiful, branching. | ||||||||
| pileus context | from RET study of paratypes: filamentous undifferentiated hyphae ?? ; acrophysalides ?? ; vascular hyphae 2.5 - 7.2 (-12.1) μm wide, rather (unusually) common especially near pileipellis, rather frequently branching, strongly sinuous. | ||||||||
| lamella trama | from RET study of paratypes: bilateral, divergent; wcs = 75± μm wide (rehydration good to excellent); with central stratum comprising interwoven filamentous undifferentiated hyphae with some intercalary inflated segments; with subhymenial base exhibiting moderate to poor rehydration, comprising plentiful fusiform to clavate cells (e.g., 62 - 69 × 17.1 - 19.0 μm), very common filamentous undifferentiated hyphae (5.0 - 6.6 μm wide, frequently branching, often constricted at septa), very common vascular hyphae in tangles (3.7 - 8.4 μm wide, with diameter inconstant). [Note: Whether the inflated cells of the subhymenial base are terminal or intercalary was not recorded in RET's notes.—ed.] | ||||||||
| hymenial trama | from protolog: trama bilateral, of the genus | ||||||||
| subhymenium |
from protolog:subhymenial tree subcellular but with diverse elongated elements from RET study of paratypes: for the most part pseudoparenchymatous (cellular), with 1 - 2 (-3) cell layers observed below longest basidia, but with some uninflated hyphal segments (some branching) found in other regions; inflated cells mostly subglobose, with basidia arising from all cells forms, with underlying part of subhymenial tree comprising short hyphal segements perpendicular to hymenial surface. | ||||||||
| basidia |
from protolog:50-60 × 14-16.5µm, tetraspored, clavate, clampless from RET study of paratypes: ca. 74 × 15.0 μm. | ||||||||
| universal veil |
from protolog: filamentous texture dominating, composed of interwoven hyphae 3-9 µm wide, diffuse sphaerocytes but not dominant, 20-50 µm wide, however vascular hyphae are numerous, clamps absent. from RET study of paratypes: On pileus surface: ??. On stipe base, exterior surface: ??. On stipe base, interior: filamentous undifferentiated hyphae 1.9 - 9.7 μm wide (commonly exceeding 9.0 μm wide), most plentiful near surfaces, branching, singly or in fascicles, interwove in open lattice, sometimes with yellow subrefractive walls, dominating except in clusters of inflated cells, occasionally constricted at septa, rather often with tip cells slightly inflated; inflated cells moderately common, clustered locally, terminal singly, broadly clavate to ellipsoid (e.g. 62 × 34 μm) or clavate (e.g., 41 - 55 × 17 - 20 μm, sometimes with walls up to 0.5 μm thick) or narrowly ellipsoid (e.g. 23 - 40 × 18.5 - 25, sometimes with walls slightly thickened), or ellipsoid (sometimes with walls up to 0.7 μm thick) or broadly fusiform or subpyriform (e.g., 21 × 15 μm, with thin walls); vascular hyphae not observed except adjacent to inner surface layer; clamps not observed. On stipe base, inner surface: only with scattered remains here and there, very thin; filamentous undifferentiated hyphae, partially gelatinized to gelatinized, predominantly sublongitudinally arranged; vascular hyphae 2.6 - 6.3 μm wide, common, coiling, sinuous, twisting, branching. | ||||||||
| stipe context | from RET study of paratypes: longitudinally acrophysalidic: filamentous undifferentiated hyphae ??; acrophysalides 137 - 320 × 25 - 39 μm; vascular hyphae ??. | ||||||||
| partial veil | absent. | ||||||||
| lamella edge tissue |
from protolog: edge cells frequent, trivial, clavate to sphaeropedunculate, width 18-27 (37.5) µm, thin-walled. from RET study of paratypes: sterile; largely collapsed and gelatinizing, in one specimen as five or more layers of inflated cells in some areas; inflated cells up to 47 × 38 μm. | ||||||||
| basidiospores |
from protolog: 9.1-12.8 × 8.3-11.4 µm, hyaline, inamyloid, subglobose or globose, monoguttulate, evident apiculus. from RET study of paratypes: [73/3/2] (9.0-) 9.1 - 12.8 (-16.5) × (8.0-) 8.3 - 11.4 (-13.5) μm, (L = 10.5 - 12.0; L' = 11.3 μm; W = 9.1 - 10.9 μm; W' = 10.1 μm; Q = (1.05-) 1.07 - 1.20 (-1.41); Q = 1.10 - 1.15; Q' = 1.12), hyaline, colorless, smooth, thin-walled, subglobose to broadly ellipsoid, rarely ellipsoid, adaxially flattened, uncommonly with "giant" spores; apiculus sublateral, cylindric; contents mono- to multiguttulate; white in deposit. | ||||||||
| ecology |
from protolog: Gregarious. From above 1000 m elev. In montane plantation of Abies cephalonica L. Rare, localized, known only from Sardinia. RET: At 1300 m elev. Associated with pure plantation of Abies cephalonica. Contu's notes on the 16-21.vii.1996 collection state, "The sole Amanita species occurring in Abies plantations in Sardinia; typically it grows only during summer together with Laccaria impolita and some Russula species." | ||||||||
| material examined |
from protolog: ITALY: SARDINIA—Prov. Sassari - Tempio Pausania, Massicio del Limbara, Punta Balistreri [1000+ m], 21.v.1994 M. Consut s.n. (paratype, CAG), 14.vi.1994 M. Contu s.n. (paratype, CAG), 23.vi.1994 M. Contu s.n. (paratype, CAG), 21.vi.1996 M. Contu s.n. (holotype, CAG), 30.vi.1996 M. Contu s.n. (paratype, CAG), ?6.vii.1996 M. Contu s.n. (paratype, CAG), 21.vii.1996 M. Contu s.n. (paratype, CAG; paratype, RET 260-4), 21.vi.1997 M. Contus s.n. (paratype, CAG), 9.vii.1997 (paratype, CAG, labeled as "holotype" in typographical error). [Note: The paratype dated "6.vii.1996" may represent the same collection from which duplicate material marked "16.vii.1996" was sent to RET for review prior to publication of the species. RET from study of paratypes: ITALY: SARDINIA—Prov. Sassari - Tempio Pausania, Massicio del Limbara, Punta Balistreri [1300 m], 16.vii.1996 M. Contu s.n. (paratype, CAG, n.v.; paratype, RET 260-3), 21.vii.1996 M. Contu s.n. (paratype, CAG, n.v.; paratype, RET 260-4). [Note: With regard to the collection dated "16.vii.1996" see note on the material examined as listed in the protolog.—ed.] | ||||||||
| discussion |
In the protolog of this species, two different collections are listed as the type. The collection specified as the type in the Latin diagnosis is taken to be the intended holotype. This collection is listed immediately preceding the collection apparently mis-labeled as "typus" in the list of materials examined. We believe it is reasonable to consider the latter reference to the "typus" as a typographical error. In RET's provisional systematics of sect. Vaginatae, the present species falls within the limits of the provisional stirps Crocea. A sporograph comparison of the taxa assigned to that stirps follows: ??more?? | ||||||||
| citations | —R. E. Tulloss | ||||||||
| editors | RET | ||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.