


| name | Amanita grossa |
| name status | nomen acceptum |
| author | (Berk.) Sacc. |
| english name | "Tasmanian Thick-Stemmed Lepidella" |
| intro | The following description is based on that of Bas (1969). |
| cap | The cap of Amanita grossa is about 100 - 140 mm wide, convex to plano-convex, soon with flattened or somewhat depressed center, fleshy, white, with nonsulcate, appendiculate margin. The cap is scattered with thin, white, irregularly shaped, crust- to patch-like, rarely wart-like, vaguely delimited, subfelted-subpulverulent remnants of volva. |
| gills | The gills are adnexed to adnate, very broad, and probably white. |
| stem | The stem is about 75 - 100 × 20 - 30 mm, equal or slightly tapering upward, solid, white, exannulate, with some thin, subfelted patches of volva on a transitional zone between the stem and the bulb. |
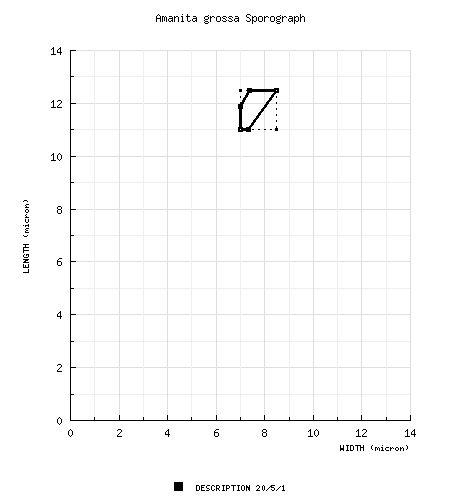
| spores | The spores measure 11 - 12.5 (-13.5) × (6.5-) 7 - 8.5 µm and are amyloid and ellipsoid to elongate. Clamps are present at bases of basidia. |
| discussion |
Amanita grossa was originally described from Tasmania. Bas (1969) knew it only from the type, but considered that it might occur on the mainland of Australia. Bas (1969) defined his stirps Grossa based on the present taxon. This stirps also includes A. ananaeceps (Berk.) Sacc., A. farinacea (Cooke & Massee) Cleland & Cheel, A. ochroterrea Gentilli ex Bas, and A. subalbida Cleland. Recently described species that can be added to this stirps with some confidence include the unusal greenish species, Amanita austroviridis O. K. Mill. and A. chlorophylla A. E. Wood. All the taxa of stirps Grossa appear to be of Gondwanan origin.—R. E. Tulloss |
| brief editors | RET |
| name | Amanita grossa | ||||||||
| author | (Berk.) Sacc. 1887. Syll. Fung. 5: 15. | ||||||||
| name status | nomen acceptum | ||||||||
| english name | "Tasmanian Thick-Stemmed Lepidella" | ||||||||
| synonyms |
≡Agaricus (Amanita) grossus Berk. in Hook. f. nom. inval. 1860. The Botany of the Antarctic Voyage. Part III. Fl. Tasmania 2: 242. [Posterior homonym. ]
non Agaricus grossus Léveillé. 1843. ??Memoire sur le genre Sclerotium. ??Ann. Sci. Nat. Bot., Sér. 2, 20: ??.
≡Aspidella grossa (Berk. in Hook. f.) E.-J. Gilbert. 1940. Iconogr. Mycol. (Milan) 27, suppl. (1): 79, tab. 51 (figs. 3-4).
??Check treatment of the latter on mycobank.org?? The editors of this site owe a great debt to Dr. Cornelis Bas whose famous cigar box files of Amanita nomenclatural information gathered over three or more decades were made available to RET for computerization and make up the lion's share of the nomenclatural information presented on this site. | ||||||||
| MycoBank nos. | 277857, 203072, 258591 | ||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||
| holotypes | K | ||||||||
| type studies | Bas. 1969. Persoonia 5: 508, figs. 292-285. | ||||||||
| revisions |
Pegler. 1965. Austral. J. Bot. 13: 334, fig. 3/9. Reid. 1980. Austral. J. Bot., Suppl. Ser. 8: 33, fig. 19(a-e), 67-68, 106. | ||||||||
| intro |
The following text may make multiple use of each data field. The field may contain magenta text presenting data from a type study and/or revision of other original material cited in the protolog of the present taxon. Macroscopic descriptions in magenta are a combination of data from the protolog and additional observations made on the exiccata during revision of the cited original material. The same field may also contain black text, which is data from a revision of the present taxon (including non-type material and/or material not cited in the protolog). Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The following text is based on the revisions of Bas (1969) and Reid (1980). Basidiomes: large, thickset. | ||||||||
| pileus |
Bas (1969): ca. 100 - 140 mm wide, white, convex to plano-convex, soon flattened or somewhat depressed over disc, smooth, "not or only very slightly polished"; context fleshy; margin nonsulcate, appendiculate; universal veil in crust- to patch-like, rarely wart-like fragments, scattered, irregularly shaped, thin, white(?), vaguely delimited, subfelted-subpulverulent, often with concentrations of crust-like and patch-like areas near disc and with crust-like zone near margin. Reid (1980): Reid viewed at least some of the "crusts" and "patches" as clusters of closely spaced small warts. | ||||||||
| lamellae | Bas (1969): adnexed to adnate, density not recorded, "very broad," up to 18 mm broad; probably white, probably with subflocculose edge; lamellulae not described. | ||||||||
| stipe | Bas (1969): ca. 75 - 100 × 20 - 30 mm, white(?), cylindric or slightly narrowing upward, fibrillose; bulb subclavate or clavate or ellipsoid or subglobose, rounded or more or less pointed [below], non-marginate, up to ca. 50 × 40 mm; context solid; exannulate; universal veil as thin patches, subfelted, in zone of transition from stipe to bulb. | ||||||||
| odor/taste | not recorded. | ||||||||
| macrochemical tests |
none recorded. | ||||||||
| pileipellis | Bas (1969): rather thick, distinctly gelatinizing near surface with age; filamentous hyphae 2.5 - 7 (-10) μm wide, interwoven; vascular hyphae present; clamps not observed. | ||||||||
| pileus context | not described. | ||||||||
| lamella trama | Bas (1969): probably bilateral. | ||||||||
| subhymenium | Bas (1969): probably ramose. | ||||||||
| basidia | Bas (1969): 4-sterigmate; clamps present. | ||||||||
| universal veil |
Bas (1969): On pileus: with elements irregularly disposed; filamentous hyphae 3 - 7 μm wide, branching rather abundant; inflated cells abundant, mainly ellipsoid or globose, sometimes pyriform or inflated-branching, 25 - 55 (-80) × 18 - 40 (-60) μm, also (close to pileipellis) elongate and up to 120 × 35 μm, terminal singly or in short chains; vascular hyphae scattered, slightly yellowish in alkaline solution; clamps not observed. On stipe: not described. Reid (1980): filamentous hyphae thin-walled, hyaline, disintegrating; inflated cells subglobose to ellipsoid, up to 35 × 25 μm, thin-walled, hyaline; "refractive oleiferous" hyphae very abundant, branched, up to 10 μm wide | ||||||||
| stipe context | Bas (1969): longitudinally acrophysalidic; filamentous hyphae 3 - 9 μm wide; acrophysalides up to 250 × 40 μm and perhaps larger; vascular hyphae present; clamps not observed. | ||||||||
| partial veil | absent. | ||||||||
| lamella edge tissue | Bas (1969): present, but impossible to rehydrate. | ||||||||
| basidiospores | Bas (1969): [20/5/1] 11.0 - 12.5 (-13.5) × (6.5-) 7.0 - 8.5 μm, (Q = (1.40-) 1.50 - 1.70; Q = 1.60), thin-walled, amyloid, ellipsoid to elongate; apiculus not described; contents not described; color in deposit not recorded. | ||||||||
| ecology | Bas (1969): Terrestrial. | ||||||||
| material examined | Bas (1969): AUSTRALIA: TASMANIA—Unkn. Municipality - unkn. loc., s.d. J. D. Hooker 1412 (holotype, K). | ||||||||
| discussion |
Bas (1969): "The type consists of several slices of fruit-bodies. In generalit is very difficult to [rehydrate] the tissues. Spores are abundant but also very difficult to [rehydrate]. "Unless pats of the volval remnants have disappeared, the volva is rather thin. In its thickest parts, at the centre of the cap, it seems as though in the inner layer of the volva, close to the pileipellis, some elongate elements are present that are lacking in the outer layer. "Among the species with clamps in subsection Solitariae, A. grossa represents a rather unusual type because of its very sturdy habit, its non-marginate, subbulbous base of the stem, and its rather thin volva, apparently consisting of irregularly disposed elements, leaving only inconspicuous remnants at the base of the stem, and scattered, small, thin patches or crusts on the cap." Bas based his stirps Grossa on the present species. The number of taxa assignable to this stirps has grown considerably with the expansion of knowledge of the Australian taxa of Amanita since 1969. It still appears that this stirps' membership is limited to Australia. [Note: Reid's description (Reid 1980) of the microscopic characters of the universal veil is more abbreviated than that of Bas and differs in stating that "refractive oleiferous hyphae" are "very abundant" rather than "scattered." The material of the type may have deteriorated in the eleven or more years between the examinations of Bas and Reid. If an observer is not cognizant of the fact that vascular hyphae are aseptate, then subrefractive or refractive septate hyphae (which may or may not be Amanita tissue) may be mistaken for vascular hyphae. In my experience, hyphae of some molds are sometimes distinctly refractive and can be confused with the vascular hyphae of an Amanita. Possible confusion of the organism represented by individual hyphae also may have led to Reid's reporting clamps on hyphae in the universal veil. In a related passage, Reid suggests that A. grossa sensu Aberdeen may be more similar to A. grossa because of his experience of finding the abundant "refractive oleiferous hyphae" mentioned above. As noted by Reid (1980), a modern revision of the present species is very much to be desired.—ed.] | ||||||||
| citations | —R. E. Tulloss | ||||||||
| editors | RET | ||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.