


| name | Amanita zambiana |
| name status | nomen acceptum |
| author | Pegler & Piearce |
| english name | "Zambian Slender Caesar" |
| images |
 1. Amanita zambiana, Copper Belt Prov., Zambia. |
| intro | The material below is based on the original description and a type study of the species and other original research by RET. |
| cap | The cap of A. zambiana is 100 - 250 (-250) mm, globose to ovoid when young, expanding to applanate with decurved margin, lacking an umbo, viscid, undecorated, with a striate margin. It is olivaceous brown over disk, progressively paler toward margin, and white at the margin. The flesh is white and up to 10 mm thick above the stem. The volva is absent. |
| gills | The gills are free and remote, very crowded, white, ventricose, and up to 15 mm broad. The short gills are truncate to rounded truncate to subattenuate, of three or more lengths, and unevenly distributed. |
| stem | The stem is 100 - 150 × 15 - 20 mm, whitish, stout, cyndrillic, fibrillose, and soon hollow. The sac-like volva is 50 - 90 × 30 - 40 mm, blackish brown on exterior surface which cracks into plaques, red-brown on inner surface in most mature exsiccata; it is pallid at base. |
| spores | The spores measure (9.9-) 10.0 - 13.5 (-21.0) × (7.0-) 7.8 - 10.8 (-12.5) µm and are broadly ellipsoid to ellipsoid (occasionally subglobose or elongate) and inamyloid. Clamps are common at bases of basidia. |
| discussion |
The species was originally described from Zambia and is common in season in central Africa. This name may be a synonym of Amanita loosii Beeli (originally described from Congo); however, the latter is described as being entirely white from the first and developing some discoloring over the center of the cap. I have examined considerable material of this African species, but have never seen it fresh.—R. E. Tulloss |
| brief editors | RET |
| name | Amanita zambiana | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| author | Pegler & Piearce. 1980. Kew Bull. 35(3): 483, 485, fig. 2. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| name status | nomen acceptum | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| english name | "Zambian Slender Caesar" | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| etymology | Zambia + -ana, suffix indicating possession; hence, "of Zambia" | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| MycoBank nos. | 112741 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| holotypes | K | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| type studies | Tulloss, here. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| selected illustrations |
Piearce. 1977. Orbit Mag. 5: 13. [n.v.] Parent & Thoen. 1977. Econ. Bot. 31: 439, fig. 8. Buyck. 1994. Ubwoba: Champ. comest. ouest Burundi&: cover, figs. 2, 54-55. [N.B.: Buyck proposes synonymy of the present species with Beeli (1936: 90, pl. III (fig. 1)) the type of which has not yet been reviewed by Tulloss.] Ryvarden et al 1994. Larger Fung. S. Centr. Africa: 78, bottom plate. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| intro |
The field may contain magenta text presenting data from a type study
and/or revision of other original material cited in the protolog of the present taxon.
Macroscopic descriptions in magenta are a combination of data from the protolog and
additional observations made on the exiccata during revision of the cited original
material. The same field may also contain black text, which is data from a revision of the present taxon (including non-type material and/or material not cited in the protolog). Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The following information is based on the protolog of the present species and original research by R. E. Tulloss. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| pileus | 100 - 200 (-250) mm wide, olivaceous brown over disk, progressively paler toward margin, white at margin, globose to ovoid when young, expanding to applanate with decurved margin, lacking an umbo, viscid, glabrous; context white, up to 10 mm thick in disk; margin sulcate-striate (0.1+R), nonappendiculate; universal veil absent. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| lamellae | free and remote, very crowded, white, ventricose, up to 15 mm broad, with edges floccose-denticulate; lamellulae truncate to rounded truncate to subattenuate, of three or more lengths, unevenly distributed. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| stipe | 100 - 150 × 15 - 20 mm, whitish, stout, cylindric, fibrillose; context fistulose, soon hollow; partial veil superior, membranous, pendulous, faintly striate above, white; universal veil as saccate volva, 50 - 90 × 30 - 40 mm, blackish brown on exterior surface, red-brown on inner surface in most mature exsiccatum, pallid at base (below surface of substrate) and in "button" specimens, apparently darkening unevenly (note dark plaques or regions in specimens of moderate age), fleshy, squamose-reticulate, with exterior layer "cracking into large, indefinite verrucae" (i.e., areolate with cracks paler than regions demarcated by them), often extending below well stipe base and often attached to sides of stipe for 1/3 to 2/3 of height of volval limb, with limbus internus at point of limb's attachment to stipe. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| odor/taste | not recorded. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| macrochemical tests |
none recorded. Market commodity, EDIBLE. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| pileipellis | 150+ - 215 µm thick, of two distinct layers; suprapellis 15 - 30 µm thick of gelatinized hyphae; subpellis yellow-brown to orange-brown, of partially gelatinized to nongelatinized hyphae; filamentous, undifferentiated hyphae 1.5 - 4.0 µm wide, branching, subradially arranged, densely packed; vascular hyphae 7.2 - 9.0 µm wide, relatively common, occasionally branched; clamps present. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| pileus context | filamentous, undifferentiated hyphae 1.2 - 12.8 µm wide, branching, plentiful, in fascicles and singly in loosely woven lattice, occasionally with yellowish subrefractive walls, with intercalary segments inflated to 36 µm wide; acrophysalides subfusiform to clavate to broadly clavate, plentiful, up to 215 × 49 µm or larger, thin-walled; vascular hyphae 3.0 - 11.8 µm wide, sinuous, scattered, may be relatively common locally, scarce in some mounts; clamps present. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| lamella trama |
bilateral, badly collapsed in holotype, immature and partially collapsed in paratype; subhymenial base and central stratum collapsed; parameters unmeasurable; filamentous, undifferentiated hyphae 1.5 - 8.2 µm wide, branching (with intercalary segments up to 25 µm wide per protolog); divergent, terminal inflated cells could not be distinguished because of condition of tissues; vascular hyphae not observed; clamps present. non-type: subhymenial base dominated by elongate curving intercalary cells (up to 122 × 14.0 µm or longer) giving rise to cells of subhymenium most distant from hymenial surface. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| subhymenium |
parameters unmeasurable because of state of tissues; pseudoparenchymatous, with two to three layers of cells below bases of longest basidia/-oles, with basidia arising from small inflated cells (and possibly other elements?). non-type: pseudoparenchymatous, with basidia arising from inflated cells and (occasionally) partially inflated cells, with 1 to 3 cells below bases of longest basidia/-oles (with these bases 10 - 15 µm distant from subhymenial base) and 4 to 6 cells below bases of shortest basidioles (with these bases up to 55 µm distant from subhymenial base), with largest cells (up to 22 × 18.9 µm) often cells most distant from hymenial surface. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| basidia |
41 - 64 × 10.8 - 14.8 µm, thin-walled, 4- or (occasionally) 1-sterigmate, with sterigmata up to 7.0 × 2.5 µm (up to 8 µm long per protolog); clamps common and prominent. non-type: 46 - 57 (-78) × 11.8 - 13.5 (-18.5) µm. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| universal veil | On pileus: absent. On stipe base, exterior surface: brown to brownish red to brownish orange, extensively collapsed and partially or strongly gelatinized in layer 45 - 60 µm thick; filamentous, undifferentiated hyphae 1.2 - 6.8 µm wide, in fascicles without dominant orientation, dominating; vascular hyphae impossible to distinguish due to state of tissue. On stipe base, interior: filamentous, undifferentiated hyphae 1.2 - 4.8 µm wide, branching, plentiful to dominant, singly and in broad fascicles forming loosely woven lattice, sometimes loosely coiled, sometimes with yellowish subrefractive walls; inflated cells ovoid to clavate to narrowly clavate, thin-walled, up to 113 × 54 µm, rather common away from surfaces; vascular hyphae 2.8 - 20 µm wide, scattered to rather frequent locally, sinuous, infrequently branching, occasionally with isolated coil; clamps rather common. On stipe base, inner surface: almost completely gelatinized, colorless at very surface for depth of 1±A± hyphal diameter, brownish yellow to brownish orange and extensively collapsed to strongly to partially gelatinized in layer 75 - 95 µm thick; filamentous, undifferentiated hyphae 1.2 - 4.2±A± µm wide, densely packed, in broad fascicles somewhat loosely interwoven and without dominant orientation, with tip cells disproportionately common at surface (in short fragments sufficiently well-preserved to have distinct walls); vascular hyphae impossible to distinguish at surface due to state of tissue. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| stipe context | longitudinally acrophysalidic; filamentous, undifferentiated hyphae 1.5 - 6.5 µm wide, branching, plentiful; acrophysalides dominant in interior, thin-walled, up to 160 × 37 µm; vascular hyphae 3.2 - 4.0 µm wide, scarce; clamps present. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| partial veil | filamentous, undifferentiated hyphae 1.0 - 7.5 µm wide, with almost all < 4.0 µm wide and many < 2.0 µm wide, branching, dominating, in fascicles or singly, at many angles, but dominated by subradially oriented fascicles tightly packed, with occasional partially inflated tip cells (e.g., 13.0 × 6.2 µm); inflated cells thin-walled, clavate, scattered, hard to distinguish, up to 33 × 13.0 µm; vascular hyphae not observed; clamps present. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| lamella edge tissue | not described. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| basidiospores |
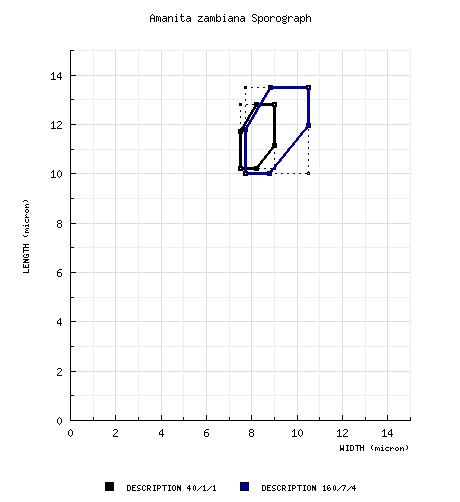
type study of RET: [40/1/1] (10.0-) 10.2 - 12.8 (-21) × (7.0-) 7.5 - 9.0 (-11.0) µm, (L = 11.5 µm; W = 8.2 µm; Q = (1.18-) 1.24 - 1.56 (-1.91); Q = 1.41), hyaline, colorless, thin-walled, smooth, inamyloid, broadly ellipsoid to ellipsoid, occasionally elongate, often adaxially flattened, occasionally with apical flattening, rarely constricted, often expanded at one end; apiculus sublateral, cylindric, prominent; contents granular or monoguttulate; color in deposit not recorded. Composite for all material revised by RET: [160/7/4] (9.9-) 10.0 - 13.5 (-21.0) × (7.0-) 7.7 - 10.5 (-12.5) µm, (L = 10.8 - 12.3 (-13.0) µm; L’ = 11.6 µm; W = 8.2 - 9.5 (-10.5) µm; W’ = 9.1 µm; Q = (1.09-) 1.14 - 1.53 (-1.91); Q = 1.19 - 1.32 (-1.41); Q’ = 1.29). | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ecology |
Solitary or in small groups. In miombo woodland, probably mycorrhizal with Brachystegia, locally abundant after the start of rains (December and January), may be found as late as March. non-type: Burundi: Gregarious at 950 m elev. Under Brachystegia utilis. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| material examined |
type study of RET: ZAMBIA: COPPERBELT PROV.—ca. Kitwe, i.1974 M. H. Ivory FP 176/66 (holotype, K [M]29729); btwn. Kitwe & Ndola, purchased at roadside, i.1975 G. D. Piearce 21 (paratype, K [M]29731). Pegler and Shah-Smith (1997): ZAMBIA: COPPERBELT PROV.—Kitwe, i.1966 M. H. Ivory 176/66 (holotype, K [M]29729); btwn. Kitwe & Ndola, purchased at roadside, i.1975 G. D. Piearce 21 (paratype, K [M]29731), 22.xii.1978 G. D. Piearce FP602/1 (K [M]29734). Mpika Distr. - North Luangwa Nat. Pk., 13.xii.1994 D. Shah-Smith 75 (K [M]29009). RET: BURUNDI: ca. Rumonge, Nyamirambo, 18.iii.1994 Assoc. des Femmes de Mutambara s.n. [B. Buyck 5529] (in herb. Buyck; RET 261-3). ZAMBIA—Copper Belt Prov. - Greystone Farm, off Kitwe-Ndola Rd., 14-24.xii.2000 David Arora 00-422 (RET 343-10). ZIMBABWE: ca. HARARE CITY—ca. Lk. McElwaine, | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| discussion |
The more mature of the two specimens of the holotype collection of the present species was apparently dried in the early stages of sporulation; occasional giant spores are present on the lamellae. Both specimens in the paratype collection are immature As a result, the data on spore size and shape is very limited and not reflective of what is to be expected in mature material. Thanks to Mr. David Arora (Mendocino, California) and Dr. Bart Buyck (PC), I have been able to obtain recent material of this species to review. In some of this material, the subhymenium is rehydratable although the subhymenial base is partially collapsed. Nevertheless, a few observations were made that expand the type study data (above). Amanita loosii appears to be somewhat similar, but is depicted in its protolog as having a volval sac attached only to the very base of the stipe and having no portion of the volva projecting noticeably below the stipe's base. The fact that most of the specimens examined were purchased in markets or from market collectors may have resulted in the spores being abnormal. Future studies are required to ascertain with certainty the size and shape of mature spores. Pegler and Piearce (1980) recorded two names for this species in languages of various peoples in Zambia, and this list was expanded by Buyck (1994) who included information gathered by Beeli (1936) on A. loosii. In Malawi: utenga (Yao), ndezu (Tonga). In Republic of Congo: futi (Kilamba), kilva (Kasengo), ntelia (Kibemba), tedia (ca. Lubumbashi, formerly Elisabethville), tente (Kiswahili). In Zambia: tente (Bemba) and ndelema (Chewa, Kibemba, Kaonde, Nyanja, Tumbuka). | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| citations | —R. E. Tulloss | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| editors | RET | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
| name | Amanita zambiana |
| bottom links |
[ Keys & Checklists ] |
| name | Amanita zambiana |
| bottom links |
[ Keys & Checklists ] |
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.